Authors Bob Thoolen Fiebo J.W.Ten Kate Domenico Castigliego Paul J.van Diest David E.Malarkey Susan A.Elmore Robert R.Maronpot
Global Pathology Support, The Hague, Netherlands, Division of Laboratories and Pharmacy, Department of Pathology, University Medical Center Utrecht, Utrecht, Netherlands, National Toxicology Program, Cellular and Molecular Pathology Branch, National Institute of Environmental Health Sciences, NC, USA, Maronpot Consulting, LLC, Raleigh, NC, USA
Previously identified comparable morphological features of human and rat hepatoproliferative lesions were identified, including hepatocellular carcinomas (HCCs). In this study we identified similarities and differences in immunohistochemical (IHC) detection of some key proteins important in carcinogenesis that may link pathogenesis pathways of human and rat HCC. The comparative features of six IHC markers (Ki-67, beta-catenin, CD34, glutamine synthetase (GS), c-myc, and transforming growth factor alpha (TGF-alpha)), previously shown to be positive or altered in rodent and human HCC when compared to normal hepatocytes, were investigated. Glutamine synthetase (a hepatocellular enzyme) was strongly positive in 5/5 (100%) human and 4/5 (80%) rat HCCs examined. CD34 (an endothelial marker) and Ki-67 (a cell proliferation marker) were consistently positive in five human HCCs (5/5 for both markers) but weakly positive in only 2/5 and 3/5 rat HCCs, respectively. Beta-catenin, c-myc, and TGF-alpha were infrequently altered, with immunopositivity found in only one or two of either rats or human HCCs (a total of five each examined; beta-catenin: 1/5 rat samples, c-myc: 2/5 human samples, TGF-alpha: 1/5 rat samples positive). It was concluded that Ki-67, CD34, and GS, are most likely to be useful in studying the pathogenesis of HCC in rats and humans.
Published online: 18 Nov 2013
Keywords
Beta-catenin, Carcinogenesis, CD34, c-myc, Glutamine synthetase, Hepatocellular carcinoma, Human, Immunohistochemistry, Ki-67, Liver tumors, Rat, TGF-alpha
Abbreviations
ABC = Avidin-Biotin Complex (ABC) method.
AFP = Alpha-fetoprotein.
BASM = Bond automated staining machine.
Beta-catenin = Cell surface glycoprotein subunit of the cadherin. Protein complex/component of Wnt-signaling pathway.
CD34 = Type I transmembrane glycoprotein that belongs to the CD34/podocalyxin family of sialomucins.
C-myc = Regulator gene/proto-oncogene.
DAB = 3,3’ diaminobenzidine tetrahydrochloride.
EDTA = Ethylenediaminetetraacetic acid.
GS = Glutamine synthetase, enzyme that catalyzes the synthesis of glutamine from glutamate and ammonia.
HCA = Hepatocellular adenoma.
HCC = Hepatocellular carcinoma.
H & E = Hematoxylin and Eosin.
HGF receptor = Hepatocyte growth factor receptor.
IHC = Immunohistochemical or immuno histochemistry.
Ki-67 = Nuclear protein associated with proliferation.
NTP = National Toxicology Program.
RT = Room temperature.
TGF-alpha = Transforming/tumor growth factor alpha.
UMCU = University Medical Center Utrecht.
Introduction
The liver is a major target organ in the safety assessment of preclinical toxicity and oncogenicity rodent studies and a primary target for drug-induced toxicity.1,2 Identification and appropriate interpretation of hepatoproliferative lesions in rodent models is critical for identifying xenobiotics and environmental factors that could potentially lead to hepatocellular carcinoma (HCC) in exposed humans. The relevance of rodent models can be further clarified by the identification of similarities and differences in proteins expressed in rodent versus human proliferative hepatocellular changes. In this study, six immunohistochemical (IHC) markers used in diagnostic human hepatopathology for the comparison of relative expression in human and rat hepatocellular carcinomas were applied.
Immunohistochemical markers for beta-catenin, glutamine synthetase (GS), Ki-67, and CD34, commonly used in human hepatopathology, have been shown to be of diagnostic value and/or changed in human or animal liver tumors, particularly in HCCs.3–16 Although less commonly used, IHC markers for c-myc17–22 and transforming growth factor alpha (TGF-alpha)19,23–26 have also shown positive expression in human and/or animal HCCs.
While human and rat HCCs are morphologically similar,27 this IHC study is an attempt to determine if commonly used IHC markers for characterizing human HCCs might also be useful in identifying similarities and differences in IHC staining between rat and human HCCs. Similar IHC responses might then be applied to further investigate the pathogenesis of rat HCC and help determine the relevance of rat hepatocarcinogenesis in safety assessment studies as predictive of human liver cancer risk.
Methods and Materials
Hepatocellular carcinoma samples
Formalin- fixed, paraffin- embedded tissues of human hepatocellular carcinomas obtained from surgical liver specimens (Department of Pathology, Division of Laboratories and Pharmacy, University Medical Center Utrecht) were used for this study. Paraffin blocks of rat hepatocellular carcinomas were obtained from the National Toxicology Program (NTP) studies of chemically induced hepatocellular neoplasms. The rat HCCs were selected as representatives of well-defined classical HCCs and were from rats exposed to tetrachloroethylene (one male F344 rat), o-nitrotoluene (two male F344 rats), tetrachloroazobenzene (one female Sprague-Dawley rat), and polychlorinated biphenyl 126/153 (one female Sprague-Dawley rat) for the 2-year carcinogenicity studies. A treatment-related induction of hepatic neoplasia was identified in these four, 2-year carcinogenicity studies. Human samples of HCCs were derived from patients with liver cirrhosis caused by Hepatitis C (one male and one female), liver hemochromatosis with severe fibrosis (one male), (non-alcoholic) steatohepatitis (one male), and one female with HCC of unknown etiology.
All neoplasms were first reviewed by a medical pathologist and two toxicologic pathologists and were confirmed as hepatocellular carcinomas based upon contemporary diagnostic criteria.28–30 The first and the last slide of each tumor specimen was stained with hematoxylin and eosin (H&E) to confirm the diagnosis and presence of adequate tumor tissue in the intervening serial sections to be used for IHC. In this pilot study, five human and five rat prototypical hepatocellular carcinomas were selected for IHC staining.
Immunohistochemical staining
Formalin- fixed, paraffin- embedded sections were deparaffinized with xylene, rehydrated, and subjected to antigen retrieval. One slide/block and five slides/markers were used for staining. Following non-specific blocking steps, polyclonal and monoclonal primary antibodies were applied according to the following method and at the following dilutions: Ki-67; clone MIB-1, mouse monoclonal anti-human MIB-1 (Dako M7240, Glastrup, Denmark) at 1:200, and clone SP6; rabbit monoclonal anti-rat (Neo-markers RM 9106-S, Fremont, USA) at 1:100, CD34; mouse monoclonal anti-human CD34 (Immunotech 0786, Marseille, France) at 1:800 and polyclonal goat anti-rat CD34 (R&D Systems AF4117, Minneapolis, USA) at 1:100, beta-catenin; monoclonal mouse anti-human/anti-rat-beta-catenin (Novocastra NCL-B-Cat, Newcastle, UK) at 1:20, GS; monoclonal mouse anti-rat/anti-human GS (BD Transduction Laboratories, Heidelberg, Germany) at 1:1600, c-myc; rabbit monoclonal anti-human/anti-mouse/anti-rat c-myc (Abcam, ab 32072, Cambridge, UK) at 1:100 and TGF-alpha; sheep polyclonal anti-rat (Lifespan Biosciences LS-C42194/32940, Seattle, USA) at 1:200. Ethylenediaminetetraacetic acid (EDTA) pre-treatment was performed for beta-catenin, GS, CD34 (anti-rat), Ki-67 (SP6 clone), c-myc, and TGF-alpha staining. For Ki-67 (MIB-1) and CD34 (anti-human), a citrate buffer was used for antigen retrieval. For all immunostains, except CD34 (anti-rat) and TGF-alpha (anti-rat), which were stained manually, staining was performed using the Bond automated staining machine (Leica, Wetzlar, Germany) with the Bond polymer refined detection kit (Leica, Cat. no DS9800, Newcastle, UK).
For CD34 and TGF-alpha the manual staining procedure was as follows: slides were incubated first with unlabeled anti/serum for 60 minutes, rinsed with PBS (3 x 3 minutes), slides were incubated with 150 µg biotin-labeled second step anti-serum for 30 minutes, rinsed with PBS/tween (3 x 3 minutes), 3,3’ diaminobenzidine tetrahydrochloride (DAB) solution was prepared, slides were rinsed with PBS (3 x 5 minutes), rinsed in phosphate/citrate buffer pH 5.8 (3 minutes), slides were developed with 200 µl DAB-reagent per slide (10 minutes), rinsed with water, and counterstained with hematoxylin according to Mayer.
In brief, the protocol sequence on the Bond stainer was antigen retrieval with epitope retrieval one and two, primary antibody for 15 minutes at room temperature (RT), and Bond polymer (second antibody) for 8 minutes at RT, DAB for 10 minutes at RT. All staining for a given protein was performed as a single batch at the University Medical Center Utrecht (UMCU) using positively charged coated glass slides (MicroSystems Leica, Bretton, UK) (for details see Table 1). Normal liver was present in the stained slides for comparison with HCC staining. Positive controls (Ki-67/MIB-1, tonsil; beta-catenin Burkitt lymphoma; CD34, tonsil; GS, perivenular hepatocytes; c-myc, colorectal carcinoma) from tissues known to exhibit positive expression of proteins of interest were used for visualization. Sections were then dehydrated in graded alcohols, cleared in xylene and cover slipped.
Background relevance of IHC markers used in this study
Ki-67 antigen is a nuclear protein, which is defined by reactivity with the monoclonal antibodies from the Ki-67 clone. The Ki-67 antigen is expressed during all active phases of the cell cycle (G1-, S-, G2-, and M- phases), but is absent in quiescent cells (G0-phase).31 Immunostaining with monoclonal antibody Ki-67 provides a reliable means for rapidly evaluating the growth fraction of normal and neoplastic cell populations. It is commonly used in diagnostic tumor pathology, e.g., in soft tissue sarcomas, prostate cancer, and breast cancer.32–34 Change in the balance between proliferation and apoptosis in the course of hepatocellular carcinoma development and progression is also known to reflect a change in the Ki-67 index. Immunohistochemical study of Ki-67, as a marker for hepatocellular proliferation, is increased in the tumor tissue as compared to normal liver tissue.11,35,36 Some investigators have correlated the Ki-67 labeling index with the clinical stage, histological grade, and as an independent prognostic indicator for patients with HCC after resection.6,37
Beta-catenin, a protein that functions in cadherin-mediated cell–cell adhesion as well as in signal transduction, has received increasing attention in recent years due to its role as an oncogene in various human cancers. Beta-catenin forms a complex with c-Met (HGF receptor) and beta-catenin, and HGF-mediated signaling pathways cooperate in hepatocyte proliferation, which may be crucial in liver development, regeneration following partial hepatectomy, and pathogenesis of hepatocellular carcinoma.38 Mutations affecting phosphorylation sites in the beta-catenin gene have been implicated in the development of human and rodent HCCs. The Wnt/beta-catenin signaling pathway can be activated in human HCCs.14,38,39 Studies suggest that nuclear translocation of beta-catenin and activation of Wingless/Wnt signaling may represent an early event in liver carcinogenesis, providing a growth advantage in a subset of hepatic tumors with a more differentiated phenotype.14,40,41 Mutations affecting phosphorylation sites in the beta-catenin gene have been implicated in the development of human and rodent HCCs.14,39,42–49 Beta-catenin activation in hepato-cellular adenomas (HCAs) is observed in men and women, and specific risk factors, such as male hormone administration or glycogenosis, are associated with their development. Immunohistochemistry studies show that these HCAs over express both beta-catenin (nuclear and cytoplasmic) and GS. This group of tumors has a higher risk of malignant transformation into hepatocellular carcinoma.50,51
CD34 is an 80-100kDa type I transmembrane glycoprotein that belongs to the CD34/podocalyxin family of sialomucins. It is a widely used marker of activated hematopoietic stem/progenitor cells and endothelium and may function as an adhesion molecule.52 It stains the endothelium of blood vessels (including alveolar wall capillaries and glomeruli, but not hepatic or splenic sinusoids).53 Sinusoidal capillarization may occur in relation to increased blood flow. These newly formed arteries and capillarized sinusoids can be stained for CD34 in contrast to normal sinusoidal endothelial cells that do not express CD34.54 Angiogenesis plays an important role in tumor growth and metastasis during hepato-carcinogenesis, and CD34 positivity is increased, especially in moderately to poorly differentiated HCCs. CD34 labeling index is considered an independent risk factor for the development of HCC and may have diagnostic value in the progression and in distinguishing HCC from non-HCC.6,8,10,55,56
Glutamine synthetase catalyzes the synthesis of glutamine from glutamate and ammonia. The expression in normal liver of mice and humans is restricted to a small population (<8%) of hepatocytes that surround the terminal hepatic venules.57,58 It is upregulated in many human HCCs and is shown to be positive in mouse liver adenomas and carcinomas.58,59 Glutamine synthetase immunopositivity, in combination with beta-catenin mutations, is an important feature of HCC in both humans and mice.40,43,45–49 In addition, some have claimed that GS may be a novel serum marker for early HCC, especially for those patients with low alpha-fetoprotein levels (≤200 ng/ml).4
Amplification of c-myc has been implicated in the pathogenesis and progression of several human neoplastic diseases, and c-myc amplification in HCC has been indicated as an unfavorable prognostic indicator.18 The c-myc proto-oncogene is frequently deregulated in liver tumors. Furthermore, enforced expression of c-myc in the liver promotes the development of hepatocellular carcinomas, a process that is accelerated by co-expression with TGF-alpha.19 Since c-myc is a known inducer of wild type p53, decreased c-myc expression may lead to uncontrolled cell growth because of the lack of p53 expression that normally induces apoptosis.60 The c-myc gene has also been shown to be expressed in the adjacent liver tissue and in some of the human cirrhotic livers. However, considerable evidence exists to prove that the expression of c-myc gene (proto-oncogene) is involved in the process of human hepatocarcinogenesis.61 The c-myc gene amplification and the consequent overexpression of the amplified oncogene have been detected frequently in various types of solid tumors, including HCC, and may play an important role in tumor pathogenesis, probably because the overexpression of the oncogene confers a growth advantage.
Transforming growth factor-alpha (TGF-alpha) is a mitogenic growth factor that plays a role in the development of liver cancer in humans and rodents and increased expression of TGF-alpha, along with epidermal growth factor receptor (EGFR), occurs frequently in human hepatocellular carcinoma and may be indicative of the early stages of human hepatocarcinogenesis.26,35,62 Immunohistochemical staining cannot detect TGF-alpha and EGFR in normal livers.63 Immunohistochemical studies in rats and mice have demonstrated TGF-alpha nuclear staining of HCCs, and TGF-alpha may favor tumor progression in transgenic mice.24,64–66
Scoring IHC results
The semi-quantative scoring method per slide was based on the following: marginal=1 (0–5%), slight=2 (6–20%), moderate=3 (21–40%), marked=4 (41–80%), and severe=5 (81–100%). The scoring method is as described for the grading of liver lesions,30 with the exception that, when focal or patchy distribution of markers staining occurred instead of diffuse distribution, the highest score was considered leading. Localization was characterized as either membranous (CD34), cytoplasmic (GS, TGF-alpha), and/or nuclear (beta-catenin, c-myc, Ki-67). Immunohistochemical assessment of the proportion of cells for the staining for the nuclear antigen Ki-67 (both MIB-1 and SP6 clone) was based on the counts in three randomly selected high-power (x40 objective) fields (HPFs). In order to analyze this, slides were examined using a Zeiss AxioPlan imaging microscope coupled to a digital camera system (AxioVision) at a magnification of x400. Cells with questionable nuclear staining or those other than hepatocytes (e.g., inflammatory cells) were discounted. Necrotic, thick, and inflamed areas as well as overlapping tumor cells were avoided during evaluation. Ki-67 labeling (fraction of proliferating cells) was calculated as a mean fraction of Ki-67 positive cells divided by the total cell count for three HPFs.
Results
Immunohistochemical results
Histomorphological features of the rat and human HCC selected for IHC staining are presented in Table 2 (A, B). The rat HCCs selected for this study were expansile and the infiltrative lesions consisted of eosinophilic to amphophilic, moderately well-differentiated hepatocytes. These HCCs were multinodular with trabecular and solid growth patterns. One rat HCC had areas of necrosis and hemorrhage. Normal pre-existing liver parenchyma was present at the periphery of all rat HCCs. The human HCCs consisted of nodules of hepatocytes arranged in solid sheets with irregular trabeculae compressed by adjacent cirrhotic liver parenchyma. The human HCCs ranged from poorly differentiated to highly differentiated.
In Table 2 (A, B), the localization, number of staining features, and IHC staining scores are presented for both human and rat HCCs. All five human HCCs were immunopositive, with three of the six markers (Ki-67, CD34, and GS) and two of the five human HCCs strongly positive for c-myc. The most pronounced immunopositivity in rat HCCs was with GS (4/5), Ki-67 (3/5), and CD34 (2/5) markers, but with the exception of GS, the staining intensity was less in the rat HCCs. Five human HCCs were negative for beta-catenin and TGF-alpha, whereas 1/5 and 2/5 of rat HCCs were immunopositive for beta-catenin and TGF-alpha, respectively. Normal human and rat tissues stained negative for the markers used, with the exception of GS, which stained perivenular hepatocytes in the normal liver of both the human and rat tissues.
The proliferative marker Ki-67 tested in this study consisted of two clones. The MIB-1 clone was used for human HCCs, whereas the rat-specific SP6 clone was used on the rat HCCs (Fig. 1B). Both Ki-67 markers showed increased expression in the HCCs, with prominent staining of the mitoses in human and rat tumors.
CD34 immunostaining was consistently stronger in the human HCCs (Fig. 2B) as compared to the rat, which only showed weak staining for this marker in 2/5 of the HCCs. Unpaired (non-triadal) arteries and capillarized sinusoids were immunopositive for CD34 in 5/5 of the human and 2/5 of the rat HCCs. Non-triadal arteries are known to be easily detected with CD34 in HCCs, in contrast to normal sinusoidal endothelial cells which do not express CD34.54,67
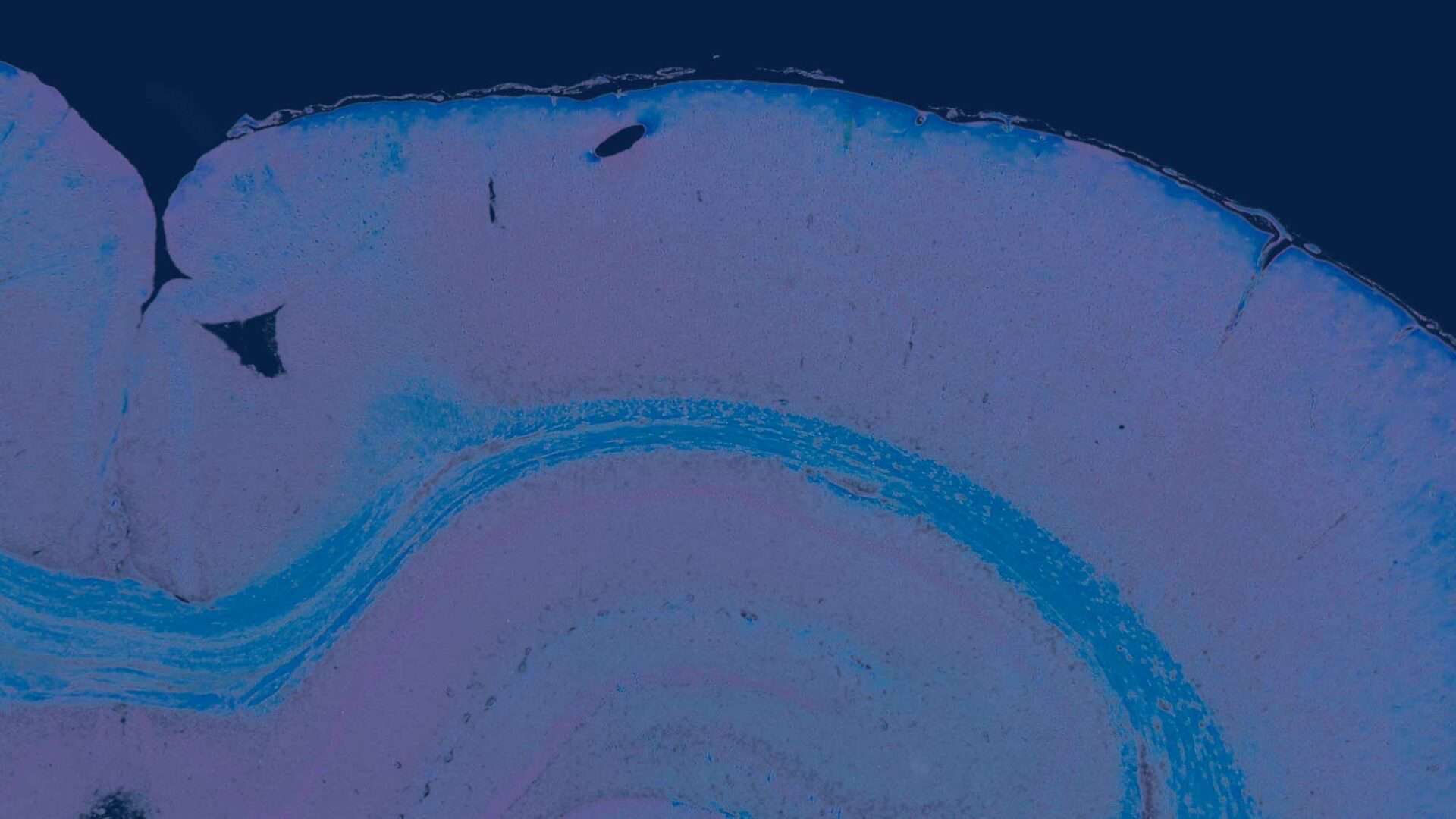
Membranous beta-catenin stain was observed in two human HCCs; however, no nuclear stain could be observed as is sometimes observed in hepatocarcinogenesis. In contrast, in the rat HCCs, one animal showed slight nuclear beta-catenin stained (Fig. 3B) in addition to membranous staining of the malignant hepatocytes.

Discussion
In preclinical testing of drugs and environmental agents, identification of carcinogens, and limitation of exposure to these substances, is one approach to reducing the human risk of hepatocarcinogenicity. Epidemiology and rodent bioassays are the predominant means by which potential human carcinogens are identified. Two classical rodent oncogenicity bioassays (rat and mouse) have been used for many years to assess the carcinogenic potential of drugs and environmental agents. In addition, it has been shown that more than half of all known human carcinogens are hepatocarcinogens in rodents.2 The use of a genetically modified mouse model, in addition to a rat, 2-year carcinogenicity study, is currently acceptable to some regulatory agencies in the conduct of preclinical safety toxicity studies.68,69 Therefore, conventional rat carcinogenicity studies continue to be a primary model for safety evaluation of new chemicals and pharmaceuticals. In these rat studies, the occurrence of spontaneous and/or xenobiotic-induced liver neoplasia such as hepatocellular carcinomas will continue to play a major role in safety assessment.
In human hepatopathology, the use of IHC markers is common practice to assist in the determination of intervention strategies and prognosis. To determine the value of these markers in rat toxicologic pathology studies, investigation of the similarities and differences between human and rat hepatocellular carcinomas based on their IHC staining patterns was performed. The six selected IHC markers were chosen based on prior publications showing their use in rodent and human hepatic neoplasia. While Ki-67, beta-catenin, CD34, and GS have been more commonly used in past studies, increased expression in human and/or rodent HCC has also been documented for c-myc21,22,70–74 and TGF-alpha.23–26,75


Beta-catenin has two distinct roles: one in E-cadherin-mediated cell adhesions, as can be observed in normal hepatocytes, and the other in carcinogenesis by activating the wnt/beta-catenin signaling pathway. One action occurs at the cell adhesion sites, where cadherins are linked to the actin-based cytoskeleton. The other action takes place in the cytoplasm and nucleus and is thought to regulate cell transformation.76 Only the nuclear staining was scored for beta-catenin because of the known role of nuclear staining in carcinogenesis.
Beta-catenin staining is involved in cadherin-mediated cell–cell adhesion as well as in signal transduction and functions as an oncogene in various human cancers.13,44–46,49 As part of the signaling of the Wnt/catenin pathway, GS immunostaining is a relevant criterion for the identification of HCC that has beta-catenin mutations and associated beta- catenin nuclear staining.4,5,9,48,77 However, in this study no nuclear beta-catenin staining was observed in the five human and rat HCCs selected, while slight to moderate GS staining was observed in all the five human liver tumors and in 4/5 of the rat HCCs. It has been reported that undifferentiated HCCs frequently have cytoplasmic/membranous beta–catenin accumulation without nuclear staining and GS expression.40 The absence of nuclear beta–catenin stain may be due to inter-individual variation in these HCCs, their degree of differentiation, and the occasional low incidence of approximately 14–34% beta-catenin positivity occurring in human HCC.40
Increased GS staining was observed in all human and rat HCCs, except for one rat HCC (animal HF691), which did not show positivity for any other IHC marker tested. The scoring for the GS staining was comparable between human and rat HCCs (Table 2; Figs. 4B and 5B).
Amplifications of the c-myc gene have been implicated in the pathogenesis and progression of several human neoplastic diseases. In addition, c-myc amplification in HCC is considered to have an unfavorable prognosis.18 In this study, c-myc staining, was only observed in 2/5 of the human HCCs, however, both had strong staining (Fig. 6B). The HCCs from these two patients also showed strong Ki-67 staining indicating the high proliferative index of these malignant tumors. None of the rat HCCs had c-myc immunopositivity.
Growth factor TGF-alpha, which plays a role in the development of liver cancer in humans and rodents, only showed marginal ‘‘patchy’’ cytoplasmic staining within the tumor of two rat HCCs.


observed (insert). The tumor is surrounded by pre-existing liver parenchyma (lower left corner) (B; beta-catenin).



The IHC staining protocol used in this study were standardized and uniformly and simultaneously applied to all human and rat HCCs to allow appropriate comparison between species and among different HCCs. For those HCCs that had variability in positive staining, or no staining, e.g., c-myc in rat HCCs or nuclear beta-catenin and TGF-alpha in human HCCs, further adjustment of antigen retrieval, antibody concentration and/or incubation times could possibly yield different results. However, in routine studies, use of standardized staining protocols is accepted practice. Consequently, at this time the reason(s) for the observed variability in IHC staining results among different HCCs cannot be explained.
Conclusion
A previous comparative study documented similar morphological features of hepatoproliferative lesions, including HCCs, in humans and rats.27 In the present investigation, IHC markers tested on human and rat HCCs revealed positive staining of some HCCs in both species. The most prominent correlation of crossspecies immunopositivity was present for GS, CD34, and Ki-67. Beta-catenin and TGF-alpha immunopositivity was present for one or two rat HCCs, respectively, and both were negative for the five human HCCs. The findings suggest that GS, CD34, and Ki-67 are most likely to be useful in studying precursor lesions involved in the pathogenesis of HCC in rats and humans.
Acknowledgements
The authors thank the following people for their technical histology and editorial support: Keith Connely (EPL, Inc.), Emily Singletary (EPL, Inc.), Mary Ellen Sutphin (EPL, Inc.), and Suzy Tirtodikromo (Global Pathology Support). Special thanks to Beth Mahler (EPL, Inc.) and Nikolas Stathonikos (UMCU) for photography support.
The majority of the photomicrographs used in this document were provided courtesy of the University Medical Center Utrecht, Department of Pathology, Division of Laboratories and Pharmacy and the National Toxicology Program Archives, National Institute of Environmental Health Sciences (NIEHS), Research Triangle Park, NC. This research was supported (in part) by the Division of the National Toxicology Program of the National Institutes of Health (NIH), National Institute of Environmental Health Sciences.
This article may be the work product of an employee or group of employees of the NIEHS, NIH, however, the statements, opinions, or conclusions contained therein do not necessarily represent the statements, opinions, or conclusions of NIEHS, NIH, or the United States government.
References
- Lee WM. Drug-induced hepatotoxicity. New Eng J Med. 2003;349:474–85. 10.1056/NEJMra021844
- Tsuda H, Futakuchi M, Fukamachi K, Shirai T, Imaida K, Fukushima S, et al. A medium-term, rapid rat bioassay model for the detection of carcinogenic potential of chemicals. Toxicol Pathol. 2010;38:182–7. 10.1177/0192623309356451
- Dal BB, Rosa L, Campanini N, Tinelli C, Torello VF, D’Ambrosio G, et al. Glutamine synthetase immunostaining correlates with pathologic features of hepatocellular carcinoma and better survival after radiofrequency thermal ablation. Clin Cancer Res. 2010;16:2157–66. 10.1158/1078-0432.CCR-09-1978
- Long J, Lang ZW, Wang HG, Wang TL, Wang BE, Liu SQ. Glutamine synthetase as an early marker for hepatocellular carcinoma based on proteomic analysis of resected small hepatocellular carcinomas. Hepatobiliary Pancreat Dis Int. 2010;9:296–305.
- Roskams T, Kojiro M. Pathology of early hepatocellular carcinoma: conventional and molecular diagnosis. Semin Liver Dis. 2010;30:17–25. 10.1055/s-0030-1247129
- Kimura H, Nakajima T, Kagawa K, Deguchi T, Kakusui M, Katagishi T, et al. Angiogenesis in hepatocellular carcinoma as evaluated by CD34 immunohistochemistry. Liver. 1998;18:14–9. 10.1111/j.1600-0676.1998.tb00121.x
- Liu L, Zhu XD, Wang WQ, Shen Y, Qin Y, Ren ZG, et al. Activation of beta-catenin by hypoxia in hepatocellular carcinoma contributes to enhanced metastatic potential and poor prognosis. Clin Cancer Res. 2010;16(10):2740–50. 10.1158/1078-0432.CCR-09-2610
- Karabork A, Kaygusuz G, Ekinci C. The best immunohistochemical panel for differentiating hepatocellular carcinoma from metastatic adenocarcinoma. Pathol Res Pract. 2010;206(8):572–7. 10.1016/j.prp.2010.03.004
- Austinat M, Dunsch R, Wittekind C, Tannapfel A, Gebhardt R, Gaunitz F. Correlation between beta-catenin mutations and expression of Wnt-signaling target genes in hepatocellular carcinoma. Mol Cancer. 2008;7:21. 10.1186/1476-4598-7-21
- Coston WM, Loera S, Lau SK, Ishizawa S, Jiang Z, Wu CL, et al. Distinction of hepatocellular carcinoma from benign hepatic mimickers using Glypican-3 and CD34 immunohisto- chemistry. Am J Surg Pathol. 2008;32(3):433–44. 10.1097/PAS.0b013e318158142f
- Pozharskyi KM, Granov DA, Ten VP, Kubaibergenova AG, Leenman EE, Rasskazov AI. The significance of immunohistochemistry in the investigation of liver neoplasms: differential diagnosis, prognostic markers. Vopronkol. 2008;54(4):417–33. 10.1179/2046023613Y.0000000026
- Shafizadeh N, Kakar S. Diagnosis of well-differentiated hepatocellular lesions: role of immunohistochemistry and other ancillary techniques. Adv Anat Pathol. 2011; 18(6):438–45. 10.1097/PAP.0b013e318234abb4
- Calvisi DF, Factor VM, Loi R, Thorgeirsson SS. Activation of beta-catenin during hepatocarcinogenesis in transgenic mouse models: relationship to phenotype and tumor grade. Cancer Res. 2001;61(5):2085–91.
- Coste de La A, Romagnolo B, Billuart P, Renard CA, Buendia MA, Soubrane O, et al. Somatic mutations of the beta-catenin gene are frequent in mouse and human hepatocellular carcinomas. Proc Natl Acad Sci USA. 1998;95(15):8847–51. 10.1073/pnas.95.15.8847
- Prange W, Breuhahn K, Fischer F, Zilkens C, Pietsch T, Petmecky K, et al. Beta-catenin accumulation in the progression of human hepatocarcinogenesis correlates with loss of E- cadherin and accumulation of p53, but not with expression of conventional WNT-1 target genes. J Pathol. 2003;201(2):250–9. 10.1002/path.1448
- Jorge A, Hashem B. Alpha-fetoprotein should be included in the hepatocellular carcinoma surveillance guidelines of the American association for the study of liver diseases. Hepatology. 2011;53(3):1060–1. 10.1002/hep.24033
- Al-Qahtani A, Al-Hazzani T, Al-hussain T, Al-Ghamdi A, Al-Mana H, Al-Arifi S, et al. Correlation between clinical characteristics, survival and genetic alterations in patients with hepatocellular carcinoma from Saudi Arabia. Cancer Genet Cytogenet. 2010;203(2):269–77. 10.1016/j.cancergencyto.2010.08.011
- Abou-Elella A, Gramlich T, Fritsch C, Gansler T. C-myc amplification in hepatocellular carcinoma predicts unfavorable prognosis. Mod Pathol. 1996;9(2):95–8.
- Cavin LG, Wang F, Factor VM, Kaur S, Venkatraman M, Thorgeirsson SS, et al. Transforming growth factor-alpha inhibits the intrinsic pathway of c-Myc-induced apoptosis through activation of nuclear factor-kappa B in murine hepatocellular carcinomas. Mol Cancer Res. 2005;3(7):403–12.
- Chandar N, Lombardi B, Locker J. C-myc Gene Amplification during Hepatocarcinogenesis by a choline-devoid diet. Proc Natl Acad Sci USA. 1989;86(8):2703–7. 10.1073/pnas.86.8.2703
- Feitelson MA. C-myc overexpression in hepatocarcinogenesis. Hum Pathol. 2004;35(11):1299–302. 10.1016/j.humpath.2004.09.012
- Peng SY, Lai PL, Hsu HC. Amplification of the c-myc gene in human hepatocellular carcinoma: biologic significance. J Formosan Med Assoc. 1993;92(10):866–70.
- Baek JY, Morris SM, Campbell J, Fausto N, Yeh MM, Grady WM. TGF-beta inactivation and TGF-alpha overexpression cooperate in an in vivo mouse model to induce hepatocellular carcinoma that recapitulates molecular features of human liver cancer. Int J Cancer. 2010;127(5):1060–71. 10.1002/ijc.25127
- Lee GH, Merlino G, Fausto N. Development of liver tumors in transforming growth factor alpha transgenic mice. Cancer Res. 1992;52(19):5162–70.
- Masui T, Nakanishi H, Inada K, Imai T, Mizoguchi Y, Yada H, et al. Highly metastatic hepatocellular carcinomas induced in male F344 rats treated with N-nitrosomorpholine in combination with other hepatocarcinogens show a high incidence of p53 gene mutations along with altered mRNA expression of tumor-related genes. Cancer Lett. 1997;112(1):33–45. https://doi.org/10.1016/S0304-3835(96)04543-0
- Moser GJ, Wolf DC, Goldsworthy TL. Quantitative relationship between transforming growth factor-alpha and hepatic focal phenotype and progression in female mouse liver. Toxicol Pathol. 1997;25(3):275–83. 10.1177/019262339702500305
- Thoolen B, ten Kate FJW, van Diest PJ, Malarkey DE, Elmore SA, Maronpot RR. Comparative histomorphological review of rat and human hepatocellular proliferative lesions. J Toxicol Pathol. 2012;23:23–36. 10.1293/tox.25.189
- Ishak KG, Goodman ZD, Stocker JT. Tumors of the liver and intrahepatic bile ducts. Third series, Fascicle 31st ed. Washington, DC: Armed Forces Institute of Pathology; 2001.
- Bannasch P, Zerban H. Tumours of the liver. In: Turusov VS, Mohr U, (eds.) Pathology of tumours in laboratory animals. 2nd ed. Lyon: IARC scientific publications No. 99; 1990. p.190–240.
- Thoolen B, Maronpot R, Harada T, Nyska A, Rousseaux C, Nolte T, et al. Proliferative and nonproliferative lesions of the rat and mouse hepatobiliary system. Toxicol Pathol. 2010;38(Suppl):5–81S. 10.1177/0192623310386499
- Gerdes J, Lemke H, Baisch H, Wacker HH, Schwab U, Stein H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J Immunol. 1984;133(4):1710–5.
- Huuhtanen RL, Blomqvist CP, Wiklund TA, Bo¨ hling TO, Virolainen MJ, Tukiainen EJ, et al. Comparison of the Ki-67 score and S-phase fraction as prognostic variables in soft-tissue sarcoma. Br J Cancer. 1999;79(5–6): 945–51. 10.1038/sj.bjc.6690151
- Borre M, Bentzen SM, Nerstrøm B, Overgaard J. Tumor cell proliferation and survival in patients with prostate cancer followed expectantly. J Urol. 1998;159(5):1609–14. 10.1097/00005392-199805000-00054
- Seshadri R, Leong AS, McCaul K, Firgaira FA, Setlur V, Horsfall DJ. Relationship between p53 gene abnormalities and other tumour characteristics in breast-cancer prognosis. Int J Cancer. 1996;69(2):135–41. 10.1002/(SICI)1097-0215(19960422)69:2<135::AID-IJC12>3.0.CO;2-8
- Daveau M, Scotte M, François A, Coulouarn C, Ros G, Tallet Y, et al. Hepatocyte growth factor, transforming growth factor alpha, and their receptors as combined markers of prognosis in hepatocellular carcinoma. Mol Carcinog. 2003;36(3):130–41. 10.1002/mc.10103
- Dutta U, Kench J, Byth K, Khan MH, Lin R, Liddle C, et al. Hepatocellular proliferation and development of hepatocellular carcinoma: a case-control study in chronic hepatitis C. Hum Pathol. 1998;29(11):1279–84. 10.1016/S0046-8177(98)90257-X
- King KL, Hwang JJ, Chau GY, Tsay SH, Chi CW, Lee TG, et al. Ki-67 expression as a prognostic marker in patients with hepatocellular carcinoma. J Gastroenterol Hepatol. 1998;13(3): 273–9. 10.1111/j.1440-1746.1998.01555.x
- Apte U, Zeng G, Muller P, Tan X, Micsenyi A, Cieply B, et al. Activation of Wnt/beta-catenin pathway during hepatocyte growth factor-induced hepatomegaly in mice. Hepatology. 2006;44(4):992–1002. 10.1002/hep.21317
- Calvisi DF, Wang C, Ho C, Ladu S, Lee SA, Mattu S, et al. Increased lipogenesis, induced by AKT-mTORC1-RPS6 Signaling, promotes development of human hepatocellular carcinoma. Gastroenterol. 2011;140(3):1071–83. 10.1053/j.gastro.2010.12.006
- Nault J-C, Zucman-Rossi J. Genetics of hepatobiliary carcinogenesis. Semin Liver Dis. 2011;31(2):173–87.
- Monga SP. Role of Wnt/β-catenin signaling in liver metabolism and cancer. Int J Biochem Cell Biol. 2011;43(7):1021–9. 10.1016/j.biocel.2009.09.001
- Hechtman JF, Raoufi M, Fiel MI, Taouli B, Facciuto M, Schiano TD, et al. Hepatocellular carcinoma arising in a pigmented telangiectatic adenoma with nuclear beta-catenin and glutamine synthetase positivity: case report and review of the literature. Am J Surg Pathol. 2011;35(6):927–32.
- Zenali MJ, Tan D, Li W, Dhingra S, Brown RE. Stemness characteristics of fibrolamellar hepatocellular carcinoma: immunohistochemical analysis with comparisons to conventional hepatocellular carcinoma. Ann Clin Lab Sci. 2010; 40(2):126–34.
- Cavard C, Terris B, Grimber G, Christa L, Audard V, Radenen-Bussiere B, et al. Overexpression of regenerating islet-derived 1 alpha and 3 alpha genes in human primary liver tumors with beta-catenin mutations. Oncogene. 2006;25(4): 599–608. 10.1038/sj.onc.1208860
- Sekine S, Lan BY, Bedolli M, Feng S, Hebrok M. Liver-specific loss of beta-catenin blocks glutamine synthesis pathway activity and cytochrome p450 expression in mice. Hepatology. 2006;43(4):817–25. 10.1002/hep.21131
- Yamaoka H, Ohtsu K, Sueda T, Yokoyama T, Hiyama E. Diagnostic and prognostic impact of beta-catenin alterations in pediatric liver tumors. Oncol Rep. 2006;15(3):551–6.
- Harada N, Oshima H, Katoh M, Tamai Y, Oshima M, Taketo MM. Hepatocarcinogenesis in mice with beta-catenin and Ha-Ras gene mutations. Cancer Res. 2004;64(1):48–54. 10.1158/0008-5472.can-03-2123
- Loeppen S, Schneider D, Gaunitz F, Gebhardt R, Kurek R, Buchmann A, et al. Overexpression of glutamine synthetase is associated with beta-catenin-mutations in mouse liver tumors during promotion of hepatocarcinogenesis by phenobarbital. Cancer Res. 2002;62(20):5685–8.
- Yamada Y, Yoshimi N, Sugie S, Suzui M, Matsunaga K, Kawabata K, et al. Beta-catenin (Ctnnb1) gene mutations in diethylnitrosamine (DEN)-induced liver tumors in male F344 rats. Jap J Cancer Res. 1999;90(8):824–8. 10.1111/j.1349-7006.1999.tb00822.x
- Bioulac-Sage P, Rebouissou S, Thomas C, Blanc JF, Saric J, Sa CA, et al. Hepatocellular adenoma subtype classification using molecular markers and immunohistochemistry. Hepat. 2007;46(3):740–8. 10.1002/hep.21743
- Bioulac-Sage P, Laumonier H, Laurent C, Zucman-Rossi J, Balabaud C. Hepatocellular adenoma: what is new in 2008. Hepatol Int. 2008;2(3):316–21.10.1007/s12072-008-9075-0
- Fina L, Molgaard HV, Robertson D, Bradley NJ, Monaghan P, Delia D, et al. Expression of the CD34 gene in vascular endothelial cells. Blood. 1990;75(12):2417–26.
- Pusztaszeri MP, Seelentag W, Bosman FT. Immunohistochemical expression of endothelial markers CD31, CD34, von Willebrand factor, and Fli-1 in normal human tissues. J Histochem Cytochem. 2006;54(4):385–95. 10.1369/jhc.4A6514.2005
- Park JO, Yeh MM. Clinical significance and implication of neo-angiogenesis in hepatocellular carcinoma. J Gastroenterol Hepatol. 2011;26(5):792–3. 10.1111/j.1440-1746.2011.06681.x
- Ohmori S, Shiraki K, Sugimoto K, Yamanaka Y, Yamaguchi Y, Saitou Y, et al. Expression of CD34-positive sinusoidal endothelial cells in patients with HBV-associated chronic liver diseases. Int J Mol Med. 2004;14(2):179–84.
- Maeda T, Adachi E, Kajiyama K, Takenaka K, Honda H, Sugimachi K, et al. CD34 expression in endothelial cells of small hepatocellular carcinoma: its correlation with tumour progression and angiographic findings. J Gastroenterol Hepatol. 1995;10(6):650–4. 10.1111/j.1440-1746.1995.tb01365.x
- Haussinger D, Schliess F. Glutamine metabolism and signaling in the liver. Front Biosci. 2007;12:371–91. 10.2741/2070
- Jeannot E, Pogribny IP, Beland FA, Rusyn I. Chronic administration of ethanol leads to an increased incidence of hepatocellular adenoma by promoting H-ras-mutated cells. Cancer Letts. 2011;301(2):161–7. 10.1016/j.canlet.2010.11.010
- Osada T, Nagashima I, Tsuno NH, Kitayama J, Nagawa H. Prognostic significance of glutamine synthetase expression in unifocal advanced hepatocellular carcinoma. J Hepatol. 2000;33(2):247–53. 10.1016/s0168-8278(00)80365-7
- Yuen MF, Wu PC, Lai VC, Lau JY, Lai CL. Expression of c-Myc, c-Fos, and c-jun in hepatocellular carcinoma. Cancer. 2001;91(1):106–12. 10.1002/1097-0142(20010101)91:1<106::aid-cncr14>3.0.co;2-2
- Zhang XK, Huang DP, Qiu DK, Chiu JF. The expression of c-myc and c-N-ras in human cirrhotic livers, hepatocellular carcinomas and liver tissue surrounding the tumors. Oncogene. 1990;5(6):909–14. 10.1159/000012024
- Schiffer E, Housset C, Cacheux W, Wendum D, Desbois-Mouthon C, Rey C, et al. Gefitinib, an EGFR inhibitor, prevents hepatocellular carcinoma development in the rat liver with cirrhosis. Hepatology. 2005;41(2):307–14. 10.1002/hep.20538
- Morimitsu Y, Hsia CC, Kojiro M, Tabor E. Nodules of less differentiated tumor within or adjacent to hepatocellular carcinoma: relative expression of transforming growth factor- alpha and its receptor in the different areas of tumor. Hum Pathol. 1995;26(10):1126–32. 10.1016/0046-8177(95)90275-9
- Vail ME, Pierce RH, Fausto N. Bcl-2 delays and alters hepatic carcinogenesis induced by transforming growth factor alpha. Cancer Res. 2001;61(2):594–601.
- Kobayashi K, Kusakabe M, Okada M, Sakairi T, Goto K, Tsuchiya T, et al. Immunohistochemical localization of transforming growth factor alpha in chemically induced rat hepatocellular carcinomas with reference to differentiation and proliferation. Toxicol Pathol. 2000;28(5):664–7. 10.1177/019262330002800505
- Sandgren EP, Luetteke NC, Qiu TH, Palmiter RD, Brinster RL, Lee DC. Transforming growth factor alpha dramatically enhances oncogene-induced carcinogenesis in transgenic mouse pancreas and liver. Mol Cell Biol. 1993;13(1):320–30. 10.1128/mcb.13.1.320-330.1993
- Roncalli M, Roz E, Coggi G, Di Rocco MG, Bossi P, Minola E, et al. The vascular profile of regenerative and dysplastic nodules of the cirrhotic liver: implications for diagnosis and classification. Hepatol. 1999;30(5):1174–8. 10.1002/hep.510300507
- Long GG, Morton D, Peters T, Short B, Skydsgaard M. Alternative mouse models for carcinogenicity assessment: industry use and issues with pathology interpretation. Toxicol Pathol. 2010;38(1):43–50. 10.1177/0192623309354107
- Storer R, Sistare F, Reddy MV, Degeorge J. An industry perspective on the utility of short-term carcinogenicity testing in transgenic mice in pharmaceutical development. Toxicol Pathol. 2010;38(1):51–61. 10.1177/0192623309351718
- Cai DW, Gao CZ, Wang NJ. C-myc gene and p53 protein expression in human primary liver carcinoma. Chin J Pathol. 1994;23(2):100–3.
- Fang YW. Expression and significance of hTERT, c-myc and Ki-67 in hepatocellular carcinoma. Cell Mol Immunol. 2009;25(4):338–40.
- Hoenerhoff MJ, Pandiri AR, Lahousse SA, Hong HH, Ton TV, Masinde T, et al. Global gene profiling of spontaneous hepatocellular carcinoma in B6C3F1 mice: similarities in the molecular landscape with human liver cancer. Toxicol Pathol. 2011;39(4):678–99. 10.1177/0192623311407213
- Pascale RM, De Miglio MR, Muroni MR, Simile MM, Daino L, Seddaiu MA, et al. C-myc amplification in pre-malignant and malignant lesions induced in rat liver by the resistant hepatocyte model. Int J Cancer. 1996;68(1):136–42. 10.1002/(SICI)1097-0215(19960927)68:1<136::AID-IJC24>3.0.CO;2-8
- Tabor E. Tumor suppressor genes, growth factor genes, and oncogenes in hepatitis B virus-associated hepatocellular carcinoma. J Med Virol. 1994;42(4):357–65. 10.1002/jmv.1890420406
- Tamano S, Merlino GT, Ward JM. Rapid development of hepatic tumors in transforming growth factor alpha transgenic mice associated with increased cell proliferation in precancerous hepatocellular lesions initiated by N-nitrosodiethylamine and promoted by phenobarbital. Carcinogenesis. 1994;15(9):1791–8. 10.1093/carcin/15.9.1791
- Wang QM, Yang KM, Zhou HY, Yu ZH, Li X, Yang HJ. Role of beta-catenin in hepatocarcinogenesis of rats. Hepatobiliary Pancreat Dis Int. 2006;5(1):85–9.
- Rignall B, Braeuning A, Buchmann A, Schwarz M. Tumor formation in liver of conditional b-catenin-deficient mice exposed to a diethylnitrosamine/phenobarbital tumor promotion regimen. Carcinogenesis. 2011;32(1):52–7. 10.1093/carcin/bgq226